UTMARK - tidsskrift for utmarksforskning
Special issue on applied ecology
http://www.utmark.org | Number 2b 2013
Peer reviewed article.
UTMARK - tidsskrift for utmarksforskning |
|
Special issue on applied ecology |
|
http://www.utmark.org | Number 2b 2013 |
|
Peer reviewed article. |
|
How does animal monitoring with GPS tags contribute to ecology and conservation?
The Global Positioning System (GPS) of the US Army was 20 years old in 2013. It has revolutionized geographical mapping and has evolved from a military survey tool to providing a broad array of public applications. Today, we can hardly imagine how to do wildlife field research without a hand-held GPS. As early as 1995, the first animals were tagged with GPS as an alternative to conventional VHF-radio telemetry. New technologies are always tempting, but have we really gained new insights in ecology by monitoring animals with GPS? Have GPS-tags actively contributed to improving management and conservation of wildlife species and wildlife-human co-existence? How can we make GPS-based research more applied? AbstractThe individual monitoring of animals with the Global Positioning System (GPS) has become common place in ecological research. To describe the contribution of this new technology to our understanding of ecology and conservation, I reviewed 174 peer-reviewed articles. While the number of articles with GPS-tagged animals has increased during the past decade, the focus of the articles has also changed gradually from methodological developments to studies on animal ecology and behaviour. In particular, the fields of animal movement, home range use and habitat selection have evolved greatly with the help of GPS. Applied research is described in 20 % of the articles included in this review and focuses primarily on the potential effects of human infrastructure on animal movement and space use. To improve the impact of GPS-based research in applied ecology, more studies should 1) use an experimental design; 2) aim to monitor a higher proportion of the population; and 3) establish the link between positioning data, behaviour and vital rates of the monitored animals. Key words: : applied ecology, behaviour, Global Positioning System, movement. Norsk sammendrag:Bruk av GPS (Global Positioning System) har blitt et vanlig verktøy i økologisk forskning. For å beskrive hvordan denne forholdsvis nye teknologien har bidratt til vår forståelse av økologi og bevaring har jeg gått gjennom 174 vitenskapelige publikasjoner. Mens antallet publikasjoner på GPS-merkede dyr har økt gjennom det siste tiåret, har fokuset forandret seg mer og mer fra metodeutvikling til studier på dyrenes økologi og atferd. Takket være GPS har studier om dyrs forflytninger, bruk av leveområder og habitatsvalg hatt spesielt stor framgang. Anvendt forskning er presentert i 20 % av de gjennomgåtte artiklene og fokuserer hovedsakelig på mulige effekter av menneskelig infrastruktur på dyrenes forflytninger og områdebruk. For å forsterke påvirkningen til GPS-basert forskning i anvendt økologi, bør flere studier 1) bruke et eksperimentelt design; 2) merke en større andel av populasjonen i fokus; og 3) lenke sammen posisjoneringsdata med dyrenes atferd og vitale mål. IntroductionFrom VHF to GPS telemetryHow can animals be monitored without disturbing them and hence altering their behaviour? This question has been intriguing many generations of biologists. Direct observations at a distance with the help of binoculars or telescopes, and ground-tracking on snow, earth or sand were the most common methods before electronic tags became available. The first radio tags emitting signal pulses at very high frequency (VHF) were attached to individuals in the early 1960s (Kenward 2001), giving rise to a new era in the study of animal ecology. VHF-signals are received by mobile or stationary receivers, and with directional antennae, the location of the animal can be estimated by means of triangulation (Rodgers 2001). This technology allowed the remote monitoring of individually marked animals to study movement behaviour, spacing, selection of habitat, and intra- and interspecific interactions. Even though VHF transmitters are relatively inexpensive, VHF tracking of animals is very time- and labour-intensive (Rodgers 2001), and the location precision using triangulation or air-tracking is relatively low (Withey, Bloxton & Marzluff 2001). The Argos satellite tracking system, based on ultra high frequency (UHF) tags, emerged in the 1980s as an alternative method of mapping long-distance movements of migrating animals. However, again, the accuracy of the positions, at several hundred meters to kilometres, is not necessarily adequate (Rodgers 2001; Bradshaw, Sims & Hays 2007). The global positioning system (GPS) went live in 1993 with the launch of the 24th Navstar satellite (Kaplan & Hegarty 2006) to orbit the Earth at a distance of roughly 20,000 km (Diggelen 2009). The system consists of three components: The orbiting satellites emitting signals continuously, ground control stations that monitor and manage the satellites, and the ground receivers that use the information received in the satellite signals to estimate their position on the globe (Kaplan & Hegarty 2006). These receiver units are commonly called GPS devices, GPS receivers, GPS units, or just simply GPS. Even though the GPS navigational system was developed by the US army and intended for military use, it soon became available for public and commercial use. Due to concerns about potential misuse, the US Ministry of Defence applied perturbations of the satellite signals for civilian use in the first years (Bhatta 2010). After disengagement of this selective availability in the year 2000, positioning precision increased from tens of meters down to just a few meters for conventional commercial GPS receivers. At the same time, the range of civilian applications developed markedly and the prices of GPS receivers dropped (Theiss, Yen & Ku 2005). As soon as GPS became available for civilian use in the 1990s, biologists started to attach GPS receivers to animals (Rempel, Rodgers & Abraham 1995; Fielitz et al. 1996; Moen et al. 1996). GPS-positions from these animal tags are stored on board, and several systems to access the positioning data while the tag is still attached to the animal have evolved (Tomkiewicz et al. 2010). Positioning data can be remotely accessed using either a satellite link (Fielitz et al. 1996; Biggs, Bennett & Fresquez 2001), a VHF- or UHF-link (Rodgers, Rempel & Abraham 1996; Rodgers 2001), or a Global system for mobile communication GSM-link (Gervasi, Brunberg & Swenson 2006; Sundell, Kojola & Hanski 2006). Because GPS tags require a lot of energy to collect and eventually transmit positions, the weight of battery packs has limited the application to large-bodied animals. An exception is the use of micro-GPS on birds and small mammals for short-term monitoring sessions of several hours to a few days, e.g. studies on homing pigeons (Meade, Biro & Guilford 2005). As battery technology is improving, applications become available for smaller animals. Battery weight is substantially reduced if the GPS-unit is solar powered. This allows GPS-applications for small-bodied animals and expands the lifetime of the GPS device (Fiedler 2009; Tomkiewicz et al. 2010). GPS tags serve as microscopes and telescopes for field ecologistsThe advantages of GPS receivers in contrast to VHF tags and UHF satellite tags are mainly the low costs combined with tracking, the pre-programming of a positioning schedule, the high frequency of positioning, and – most importantly - the high temporal and spatial precision of positioning data (Rodgers 2001). GPS accuracy after the disengagement of selective availability in year 2000 is estimated to be 22 m (Bradshaw, Sims & Hays 2007). In a similar way to the way the microscope has revolutionized our understanding of cell biology by bringing to light details that were hidden by the resolution of the human eye (Amos 2000), so GPS-tags reveal details of individual animal trajectories that cannot be assessed by direct observations or conventional radio tracking. We can describe the trajectories with step lengths and turning angles from hour to hour, minute to minute, or even from second to second for micro-GPS (Meade, Biro & Guilford 2005; Calenge, Dray & Royer-Carenzi 2009). GPS-tags also have a telescope effect: They allow us to observe animals that are beyond the reach of our eyes, either because they are too far away, because they are hidden in cover, or because darkness obscures the sight. Why this review?After the first two decades of GPS-applications in animal tracking, it is time to summarize and evaluate the current state of art. Has this new technology advanced our understanding of ecology? Has it contributed positively to the management and conservation of animal populations and wildlife-human co-existence? And what improvements are needed to make this technology more useful to science and management? This review aims to 1) describe the methodological development and challenges of GPS-based research; 2) summarize the main basic and applied ecological questions addressed in animal research with GPS-tags; 3) identify the limitations and knowledge gaps of GPS-applications; and 4) formulate recommendations on how to make GPS-based research more applied. MethodsTo find a representative sample of all literature published on animal tracking by means of GPS units, I accessed all articles in Web of KnowledgeSM (Web of Science) with the search term “GPS OR global positioning system”, refined to the Web of Science Categories “Ecology” and “Zoology” and the Web of Science Document Types “Article”, “Proceedings paper”, and “Review”. This resulted in a total of 890 hits on October 12, 2012. Because there are numerous ecological studies that used hand-held GPS to register observations in the field, I filtered the dataset in a second step by browsing through the abstracts. This left me with 522 hits. This is not a complete list of all published GPS-based research because the refinement to the specific categories “Ecology” and “Zoology” resulted in an exclusion of more general natural science journals, such as Science or the Philosophical Transactions of the Royal Society. Additionally, GPS-based animal tracking has become a standard methodology during the past decade so authors do not necessarily mention this term any longer in the search-sensitive texts, but rather in the methods chapter. I am aware that this introduces a bias into the dataset. In order to ask additional, more detailed questions from a sub-set of the total number of papers, I sorted the dataset of 522 articles by alphabetical order of the authors and read through the abstracts of the first 174 articles, corresponding to one third of all articles in my dataset. This alphabetical sampling may raise some concerns regarding repeated measures of some authors. However, of the 139 different first authors, 114 (82 %) had only one article in the database, 20 (14 %) had two, and five (4 %) were first –author of three or more articles. Repeated first authors did not necessarily publish work on the same species or topic. I categorized the content of the sampled articles into methodological, basic ecological, and applied ecological issues, with many sub-categories (Table 1). These categories were not exclusive, i.e. a specific article could cover several of the main- and sub-categories. Methodological issues covered technical aspects, data management or data analysis related to GPS-tags. If articles presented insights into the ecology of animal populations, such as migration distance or home range size, they were counted in the main category of basic ecology. If the ecological information was applied to interactions with humans, conservation or management issues, they scored in the applied ecology category. Table 1: Number of sampled articles (n = 174) for different species groups, with their content classed into the main categories: methods, basic ecology and applied ecology, and sub-categories.
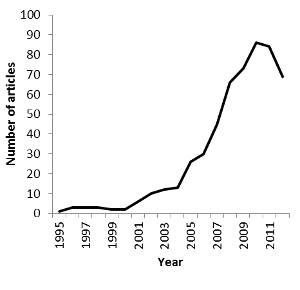
The sample of 174 articles was sufficient to look at some basic trends, such as the change of focus from methodological to ecological issues during the past two decades. When summarizing and discussing ecological applications, I have complemented the review with some selected articles that were not included in the sample. These additional articles were not included in the statistics. From methodological concerns to basic ecology to applied ecologyAmong all 522 articles found on Web of Science, the very first GPS-wildlife study to be published used GPS receivers on moose (Alces alces) (Rempel, Rodgers & Abraham 1995). After the first five-year period during which just one to three articles were published per year, the number of publications has grown exponentially and reached its maximum of 86 articles in one year, 2010 (Figure 1). The apparent decline in the last two years is probably caused by the delay of printing and the fact that the year 2012 was not complete at the time of the literature search.
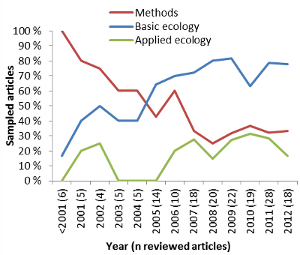
Figure 1: Number of publications identified by Web of Science using the search term GPS OR global positioning system in the Science Categories Ecology and Zoology, filtered for applications of GPS-tags on animals (n = 522). The majority of the articles within my sample were on mammals (Table 1), with 133 (76 %) on terrestrial and 5 (3 %) on aquatic mammals. Articles on birds made up 10 %. Of the three studies on reptiles (Table 1), two were on marine green turtles (Chelonia mydas) (Brooks, Harvey & Nichols 2009; Hazel 2009), and one on the terrestrial lace monitor (Varanus varius) (Flesch et al. 2009). Only one study was on a fish species (Evans et al. 2011). The application of GPS-tags to animals in aquatic environments is limited by the fact that GPS-signals quickly fade in water, and that GPS-tags can be difficult to attach without impacting the animal’s ability to swim. Fourteen articles (8 %) dealt with test tags or data simulation (Table 1). With all new technologies there is a need for development. Not surprisingly, 54 (31 %) of 174 sampled articles focused purely on methods, and additional 17 articles (10 %) put a strong emphasis on methods while at the same time presenting some ecological results (Table 1). The percentage of articles presenting methodological issues however declined during the first decade and stabilized at around 30% since 2007 (Figure 2). Focus has shifted from methodological development to ecological issues (Figure 2). In total, 120 articles (69 %) presented results on the basic ecology of animal populations, and 34 (20 %) dealt primarily with applied issues (Table 1).
Figure 2: Percentage of sampled articles (n = 174) that contain methodological, basic ecological, or applied ecological issues. The number of sampled articles per year is given in the brackets along the x-axis. Methodological considerationsOne of the main challenges when adapting GPS-technology to wildlife research is that GPS-tags have to be attached to wild animals that move in a wide variety of habitat and weather conditions, and that tags cannot easily be retrieved and checked or batteries recharged. There are several studies on the reliability and battery life span of different GPS-brands (Fielitz et al. 1996; Blake, Douglas-Hamilton & Karesh 2001; Di Orio, Callas & Schaefer 2003; Frair et al. 2004; Gau et al. 2004; Blackie 2010), the modification of GPS-tags to specific animal species, e.g. reptiles (Flesch et al. 2009; Hazel 2009), birds (Cumming & Ndlovu 2011; Guthrie et al. 2011), small mammals (Haines et al. 2006; Haramis & White 2011), and pop-up tags for pelagic fish (Evans et al. 2011), and the combination of GPS receivers with other sensors (Gervasi, Brunberg & Swenson 2006; Evans et al. 2011; Heurich et al. 2012). GPS-tags weigh more than VHF-tags due to larger battery packs. The impact a tag may have on the animal depends on the species, sex, age, body size and condition of the animal (Kenward 2000). As a rule of thumb, the weight of a tag should not exceed 3 % of the animal’s body mass of birds (Millspaugh & Marzluff 2001), and Macdonald (1978) proposed a 5 % limit for mammals. Blanc & Brelurut (1997) tested a 3.4 kg GPS-collar on tamed red deer (Cervus elaphus). Weighing 3.5 % of the total body mass of the animal, the tag may have been responsible for the observed 40% decrease in grazing activity and changes in social interactions after deployment. A slight increase in collar weight from 0.4 % to 0.6 % of the total body mass of plain zebras (Equus burchelli) may have been responsible for a 50% reduction in travel rate when zebras were foraging (Brooks, Bonyongo & Harris 2008). Half of the methodological papers focused on location errors and positioning success rates (Table 1). They described the limitations of GPS positioning in areas with limited sky view due to topography (narrow valleys, steep hill sides), vegetation with high canopy cover, or buildings in urban areas (Bowman et al. 2000; Biggs, Bennett & Fresquez 2001; D'Eon et al. 2002; Cain et al. 2005; DeCesare, Squires & Kolbe 2005). Some studies attribute positioning errors to the behavioural state of the animal, e.g. the GPS-antenna of bedded animals pointing to the ground, or animals using burrows for resting or denning (Bowman et al. 2000; Biggs, Bennett & Fresquez 2001; D'Eon & Delparte 2005). Many authors point out the bias arising from missing or erroneous positions, especially when doing habitat selection analyses (Dussault et al. 1999). Suggestions on how to correct for such biases include the simulation of missing locations based on animal trajectories (Frair et al. 2004; Etherington, Pietravalle & Cowan 2010), the application of sample weights to missing locations based on probability models (Frair et al. 2004; Horne, Garton & Sager-Fradkin 2007), and the identification of erroneous positions by establishing distance and turn-angle rules for the animal trajectory (Bjørneraas et al. 2010). The time-lag before publications about ecological issues became abundant (Figure 2) is most likely related to the handling of large datasets of highly auto-correlated spatio-temporal data (Dray, Royer-Carenzi & Calenge 2010). Researchers have had to spend a lot of time managing their datasets with conventional database programs that are often limited in space, or developing server applications for data management. Conventional movement and home range methods have been refined and new methods developed as a consequence of high frequency and more accurate positioning data (Fieberg & Kochanny 2005; Hemson et al. 2005; Dettki & Ericsson 2008; Calenge, Dray & Royer-Carenzi 2009; Bunnefeld et al. 2011; Haddadi et al. 2011). In particular, the field of resource selection modelling has developed greatly since the onset of GPS (Lele & Keim 2006; Chetkiewicz & Boyce 2009; Duchesne, Fortin & Courbin 2010). It has been proposed that autocorrelation issues can be solved with mixed models (Duchesne, Fortin & Courbin 2010) and multivariate statistics (Basille et al. 2008) in resource selection modelling, or Brownian and random bridge methods in home range utilization studies (Kranstauber et al. 2012). Basic ecologyOf the 120 papers with ecological results, more than half (53%) describe different movement parameters or use movement to describe activity patterns and foraging behaviour of the studied individuals (Table 1). The most detailed accounts of animal movement are from studies on homing pigeons (Columba livia). These birds have been fitted with micro-GPS that register positions every second during the homing flights lasting a few hours. GPS-technology has brought new insights into the importance of geo-magnetic fields for the orientation system of homing pigeons (Dennis, Rayner & Walker 2007; Gagliardo et al. 2009), and how sociality affects movement behaviour (Dell'Ariccia et al. 2008). Studies on large bird species have revealed details of the soaring behaviour during migration and foraging trips (Avery et al. 2011; Bohrer et al. 2012). GPS has also been used to study green turtle (Chelonia mydas) movement in relation to tides (Brooks, Harvey & Nichols 2009). For migrating animals, GPS-tags have been used to identify the migration route, important stop-over sites, and barriers and corridors to migration, e.g. for imperial eagles (Aquila heliaca) in Eastern Europe (Gradev et al. 2011), plain zebras in Botswana (Bartlam-Brooks, Bonyongo & Harris 2011), and savannah buffaloes (Syncerus caffer) in Western Africa (Cornelis et al. 2011). GPS-technology has definitely contributed to a better understanding of partial migration, i.e. why certain individuals of a population migrate while others do not. The sampled articles included studies on roe deer (Capreolus capreolus) (Cagnacci et al. 2011), elk (Cervus elaphus) (Hebblewhite & Merrill 2007), and moose (Bunnefeld et al. 2011). Because animals repeatedly return to and spend a considerable amount of time at foraging sites, fine scale movement data from GPS-positions have been used to reveal foraging sites, and in a second step to estimate foraging energetics. There are many studies on foraging movements of different species of sea birds, e.g. (Fritz, Said & Weimerskirch 2003; Gremillet et al. 2004; Awkerman et al. 2005; Hamer et al. 2007; Dell'Ariccia et al. 2010; Chivers et al. 2012). Foraging movements have also been studied for ungulates, (e.g. Brooks & Harris 2008; Cornelis et al. 2011). GPS-collars on large mammalian carnivores have proven useful to detect kill sites at clusters of GPS-positions and to estimate kill rates (Anderson & Lindzey 2003; Cavalcanti & Gese 2010). The social organization of animal populations is an important determinant of population dynamics. Home range size and degree of home range overlap are for example directly coupled with population density. GPS-technology allows for more precise accounts of home range size utilization. Of the 120 sampled articles with basic ecological results, 28 (23 %) presented home range estimates for the studied animals, and 17 (14%) looked at intra-specific spacing and interactions (Table 1). For example, wolverines (Gulo gulo) at their southern distribution limit roamed in relatively large home ranges that were exclusive for individuals of the same sex, and males used more than double the home range area of females (Inman et al. 2012). Jaguars (Panthera onca) in Brazil, however, are more social than previously thought, with extensive overlap of male home ranges and close encounters between individuals (Cavalcanti & Gese 2009). The most common topic of GPS-based ecological studies was habitat selection: 64 (53 %) of 120 sampled papers with ecological results described habitat or resource selection of the GPS-tagged individuals (Table 1). In contrast to the earlier VHF-based studies, the high precision of GPS-locations means that locations are now at the same spatial scale as many resource maps derived from satellite imagery and aerial photographs. This allows us to study resource selection at very fine scales and to include temporal aspects of habitat selection. The Normalized Difference Vegetation Index (NDVI) reports repeated measures of the greening of the vegetation at a very fine spatial scale and can be used as a temporally and spatially changing indicator of plant productivity. It was, for example, used to study migration routes of GPS-tagged red deer in Norway along phenological gradients (Bischof et al. 2012). Habitat selection is a multiscale process. Following Johnson’s (1980) classification, animals choose locations at the landscape level (population range, first order), within the population range at the home range level (second order), within the home range at the patch level (third order), and within the patch level at the site level (fourth order). The high spatial and temporal resolution possible is the reason why most sampled articles focused on third-order (patch level) and fourth-order (site level) selection. An example of fine-scale habitat selection is given by GPS-collared moose that used vegetation types with thermal shelter to avoid heat stress in summer (Dussault et al. 2004). Access to food is an important determinant of resource selection. However, the availability of food and water can change with season, and several studies have focused on seasonal patterns in resource selection of boreal species, e.g. woodland caribou (Rangifer tarandus) (Gustine & Parker 2008) and black bear (Ursus americanus) (Garneau et al. 2008), and savannah species, e.g. savanna buffalo (Cornelis et al. 2011). Long-term changes in habitat availability may influence population distribution and viability. Freitas et al. (2012) identified land-fast ice on Svalbard as an important habitat for GPS-collared female polar bears (Ursus maritimus) with cubs of the year, probably due to better access to more predictable prey. Climate warming is causing this habitat type to decline (Freitas et al. 2012). An interesting habitat selection study looking at the first (landscape) and second order (home range) scale (Johnson 1980) has been done on moose (Dussault et al. 2005): The authors found support for the hypothesis that the importance of limiting factors (predation > food availability > snow) was coupled with the spatial scale of habitat selection. At the landscape scale, they observed spatial segregation of wolves (Canis lupus) and moose, with moose being mainly distributed in areas with fewer wolves or with high snow cover, resulting in lower predation risk. At the home range scale, food availability and low snow cover were more important. Predation risk varies across the landscape, depending on the density and distribution of predators and the availability of shelter habitat. Studies on this so-called landscape of fear and how predation risk alters the habitat selection of prey species have become possible with the use of GPS. The group size of bison (Bison bison) was larger on meadows than in forests, and the larger the group, the more time bison spent grazing on meadows (Fortin et al. 2009). The authors link this fusion-fission behaviour of bison to predation risk. If predation risk is an important determinant of herbivore habitat selection, one might expect a trophic cascade due to a re-distribution of ungulates and subsequently changes in grazing or browsing pressure. The most well-known and discussed GPS-based studies are on how the re-colonizing wolf population in Yellowstone National Park shapes the movement of their main prey, elk, away from aspen stands, which in turn are recovering from high browsing pressure (Creel et al. 2005; Fortin et al. 2005). Studies on species interactions where individuals of at least two species have been GPS-tagged and simultaneously monitored are still rare. A study on white-tailed deer (Odocoileus virginianus) and wolves estimated that each deer home range was visited on average every 3-5 days by a wolf during summer (Demma, Barber-Meyer & Mech 2007). Wolves visited home ranges of older, more productive female while-tailed deer more often than those of two-year old females (Demma & Mech 2009). In a similar study on wolves and moose, the encounter frequency for particular moose individuals was low (Eriksen et al. 2009) and the activity pattern of moose and wolf was not correlated (Eriksen et al. 2011). The authors attribute this to the high moose densities and the large size of the wolf territory, resulting in a high prey-predator ratio. GPS-technology is also suitable for studying niche overlap and interspecific competition between species at the same trophic level. GPS-collared white-tailed deer and cattle showed space use overlap in the most productive areas but kept segregated in other areas, mostly due to different habitat requirements (Cooper et al. 2008). Even plant ecologists are using GPS when they want to find out how far seeds are dispersed by animals. GPS-tagged Asian elephants (Elephas maximus) on Sri Lanka and Myanmar carried 50% of ingested seeds over 1.2 km and are therefore an important factor for seed dispersal (Campos-Arceiz et al. 2008). The importance of animal-borne seed-dispersal is also described for GPS-tagged African elephants (Loxodonta africana) and Ramphastos toucans (Ramphastos vitellinus) in South America (Blake et al. 2009; Kays et al. 2011). Applied research based on GPS tagsOf the 174 sampled articles, 34 (20 %) described studies that were designed primarily to answer applied questions (Table 1). Another 21 (11 %) articles presented basic ecology of the GPS-monitored animals together with a conclusion on how these findings impact on the management or conservation of the population or ecosystem in study (secondary applied). Among the primary applied articles, the impact of human-made habitat alterations on natural animal populations was the most studied subject with 74% of all primary applied articles (Table 1). Due to the new insight into small-scale movement behaviour, most of these studies (20 of 26 on habitat alterations) focus on human infrastructure. Roads fragment the landscape. In order to support connectivity in natural populations, managers and landscape planners need to identify road strips with corridor habitat. Ecologists have an important role in this process by using resource selection models and movement path analyses for the species in focus. In a study of GPS-collared brown bears (Ursus arctos) and cougars (Puma concolor) in Alberta, Canada, the authors made specific recommendations on where to plan for habitat corridors and highway crossing zones for these two large, wide-roaming carnivore species (Chetkiewicz & Boyce 2009). Similarly, a study of jaguars in the Mayan Forests of Mexico and Guatemala pointed out a strip of road that was most suitable as a wildlife crossing zone for this species (Colchero et al. 2011). A study of elk in Arizona, USA, described the passage rate, i.e. crossings per approach, of a reconstructed, widened highway which was half that of the rate before or during reconstruction (Dodd et al. 2007). The authors identified stretches where fences to funnel movements to highway underpasses were most appropriate (Dodd et al. 2007). A combination of data from GPS-marked moose and moose-vehicle collisions were used to identify stretches for different mitigation measures in Quebec, Canada (Dussault et al. 2007). Once mitigation measures to increase connectivity along roads and reduce wildlife-vehicle collisions have been implemented, the effectiveness of these measures can again be tested by monitoring GPS-tagged animals. Salt pools at the road sides resulting from de-icing treatment can attract ungulates. Studies on GPS-marked moose in Quebec looked at the effect of salt pool removal and the establishment of artificial salt pools at some distance from the road (Grosman et al. 2009; Grosman et al. 2011). Rapid gas and oil development with enormous habitat alterations over large areas of Canada has triggered many studies of the impacts on animal populations. GPS-based research revealed that woodland caribou seem to be especially sensitive to human disturbance and the construction of roads, seismic lines and pipelines from industrial development (Dyer et al. 2001; Dyer et al. 2002). Pronghorns (Antilocapra americana) have also been shown to abandon areas of high industrial development in Wyoming, USA (Beckmann et al. 2012). All studies of the effects of human disturbance on wildlife in this review looked at animal behaviour in relation to habitats or times of day with high human activity. The use of GPS-tags also allows direct studies of human disturbance, by, for example, measuring the reaction of animals provoked by the approach of humans. GPS-tagged moose approached on back-country skis responded with a strong short-term increase in movement rate and enlarged daily activity range, resulting in the moose leaving the original site (Neumann, Ericsson & Dettki 2010). Although the authors consider single disturbance events as non-significant for the overall energy budget of this large-bodied species, they were concerned about repeated and intentional disturbance of wildlife by, for example, tracking animals on snow. The conservation of large animal species that show wide-ranging movements and have a high monetary value is a challenge when designing conservation areas. In Kenya, smallholder farmlands are crop-raided by African elephants that move out of conservation areas, leading to culling and poaching of elephants by the local people (Graham et al. 2009). GPS-tagged elephants seem to have adapted to the risk of predation by humans by entering the smallholder farmlands mostly by night, contrary to the more diurnal activity pattern common in protected zones and areas of higher human acceptance (Graham et al. 2009). The authors suggest establishing refuge areas, stopping the on-going habitat fragmentation by small-scale farming, and implementing and testing conflict mitigation measures in already fragmented landscapes. Poaching was the cause of death for two of seven GPS-tagged African elephants in Tanzania (Galanti et al. 2006). This article concluded with the importance of a management regime across National Park boundaries, protection of migration corridors, and the stimulation of alternative, sustainable land use practices to enhance the livelihood of local communities. The most extreme conservation measure is to completely separate animals from human risks. Several conservation areas in Southern Africa are surrounded by fences in order to keep settlers and poachers out and wildlife inside. Are these areas large enough to sustain natural populations and allow for natural movement and foraging behaviour? Re-introduced GPS-tagged lions (Panthera leo), spotted hyenas (Crocuta crocuta) and a leopard (Panthera pardus) in the Addo Elephant National Park in South Africa had home range sizes and movement activities comparable to those of animals in non-fenced areas, and the authors conclude that the fences did not affect the natural behaviour of the predators (Hayward et al. 2009). In this last example, GPS-tags were not only used to look at the effect of the fences, but also to monitor the translocated animals in order to assess the success of the re-introduction. The Chimpanzee Conservation Center in Guinea released a group of wild-born, fostered chimpanzee (Simia troglodytes) in a National Park to reinforce the natural population (Humle et al. 2011). Eight of twelve animals were GPS-tagged, and the GPS-data from retrieved collars, together with direct observations, showed that the re-integration was successful for nine animals two years after release. Detailed data on habitat selection can be used for population control in the case of pest species. Feral cats are a major threat to many natural populations, and the identification of places to set up traps can therefore help managers to control these populations (Bengsen, Butler & Masters 2012). Introduced Australian brushtail possums (Trichosurus vulpecula) are regarded as a pest in New Zealand (Blackie, Russell & Clout 2011). In a GPS-based study of the spacing behaviour of juvenile brushtail possums the authors found increased philopatry if mothers were removed, and they conclude that selective removal of problem animals will not necessarily result in a reduction in population size. In Scandinavia, managers are applying the Judas technique to limit the establishment and spread of raccoon dogs (Nyctereutes procyonoides): Sterilized raccoon dogs are tagged with GPS and followed to find and control dens of new immigrants (Dahl, Åhlén & Granström 2010). Another important applied topic is the documentation of livestock depredation by large carnivores and testing of loss-reducing mitigation measures. More than 30% of prey killed by jaguars in a study in Brazil were cattle (Cavalcanti & Gese 2010), and another study showed that male jaguars of the Yucatan Peninsula (Mexico, Guatemala) lingered more in agricultural areas and killed more cattle than females (Conde et al. 2010). A study of GPS-tagged wolves in Southern Alberta, Canada, revealed that wolves preferred to prey on livestock during the grazing season and frequently scavenged on livestock carcasses at carcass dumping places during the non-grazing season (Morehouse & Boyce 2011). The authors recommend a different carcass disposal system and livestock protection measures to make wolves less accustomed to livestock in order to decrease conflicts. Animals can be vectors of diseases that can either threaten natural populations, livestock or humans. A habitat model of mule deer (Odocoileus hemionus) and white-tailed deer in Alberta, Canada, revealed that the contact rate between animals and therefore the risk of transmitting chronic wasting disease (CWD) was density-dependent, and that the density effect was stronger in heavily wooded areas (Habib et al. 2011). In Scandinavia, managers are not only concerned about the effects invasive raccoon dogs may have as predators of native prey species, but also the potential of this species to spread rabies and parasites harmful to humans, e.g. the tapeworm Echinococcus multilocularis (Dahl, Åhlén & Granström 2010). The GPS-based Judas-technique described above is used to keep unwanted diseases and parasites in check. Conclusions and recommendations: How to make GPS-based research more appliedThe progression of GPS-based wildlife research is common to that of all new technologies in science: the method was established first, while the questions were asked later (Amos 2000). Researchers have gained access to high-resolution data on animal movement, and expectations to address important basic and applied questions in ecology are high. There are many limitations to and unused possibilities of GPS-derived data that need some consideration. Observational versus experimental studiesCause-and-effect studies are essential to the application of science, but these studies can only be achieved by experimental design. Observational studies may in the best case reveal correlations, but not effects. To base management decisions on pure correlations can be misleading and may even harm humans and conservation. Do GPS-tags contribute to a higher proportion of experimental studies in field ecology, a branch of science that mainly uses observational methods? Of the 120 sampled studies with basic and/or applied ecological results, 118 (98%) were purely observational. Only two used an experimental design. Researchers tested orientation abilities of homing pigeons with magnet-induced clock-shifting (Gagliardo et al. 2009). To test the effect of population control, another research team compared the space use and dispersal behaviour of juvenile brushtail possums before and after their mothers had been removed (Blackie, Russell & Clout 2011). Of the 118 observational studies, 97 (82%) were purely descriptive, 13 (11%) were comparative, and 8 (7%) used GPS-data to model effects of different sampling frequencies or analysis methods. More studies using experimental design are needed, especially in applied ecological projects. Sample size and sampling frequencyIn GPS-studies, the sampling unit is the individual animal, with GPS-positions being repeated measures of the same individual. The sample size refers therefore to the number of tagged individuals, whereas the sampling frequency refers to the number of GPS positions per individual. Inferences of individual-based studies at the population level are necessary if management or conservation is supposed to act on populations. For a sample of animals to be representative of a population, a sufficiently large number of animals representing an unbiased proportion of the population has to be monitored. GPS tags are however expensive, and researchers can rarely afford to mark enough animals to be able to make inferences at the population level (D'Eon & Serrouya 2005; Girard et al. 2006; Hebblewhite & Haydon 2010). Of the 96 papers with information about sample size in the abstract, 44 (46 %) report sample sizes of ten or less animals. The median number of monitored individuals is 11.5, and only 10 papers (10 %) present sample sizes > 50 individuals. A typical bias of GPS-based studies is that data are only obtained from individuals of a specific age class or sex, although sex and age can be important determinants of habitat selection, prey choice and other behaviour. Among the sampled articles, the GPS-study with the highest sample size (n = 125 female pronghorn) modelled the impact of gas field development on the habitat use of females only (Beckmann et al. 2012). From the small and often biased sample of tagged individuals, researchers fill up servers with thousands of high-resolution positions. The possibility of increasing the sampling frequency has been tempting for many scientists who now are stuck with loads of highly correlated data that are difficult to manage and analyse. It is important to bear in mind that a high sampling frequency cannot correct for low sample sizes. It is also important to ask, how much more detail is needed to describe the ecology of an animal in order to contribute essentially to conservation and management issues. From movement to behaviourWhether to move or not is usually an intentional decision of an individual, based on an array of needs and circumstances, and it is connected to different types of behaviour. But movement alone does not describe the behaviour. We can only make guesses of what an animal has been doing at place x at time y, when looking at a series of GPS positions. If not combined with biosensors that measure for example heart rate or activity (Heurich et al. 2012), with a video camera, or with field surveys of visited locations (Anderson & Lindzey 2003), we can hardly use the movement data to deduce behaviour. We can for example identify a highly selected habitat type if an animal spent a disproportionately high amount of time in this habitat, compared to its availability. We can however not conclude that this habitat type is crucial. Other less-visited habitat types may be more important for a specific, strongly fitness-related behaviour (Cooper & Millspaugh 2001). From behavioural ecology to population dynamicsIndividual fitness is a consequence of behaviour and therefore, behaviour is an important driver of population dynamics. Greater insight into the behavioural ecology will therefore improve the understanding and prediction of population dynamics (Morales et al. 2010). But do GPS-data tell us enough to make implications about fitness? Most GPS-studies monitor individuals for only a small fraction of their life history due to technical or financial limitations. Positioning data can show us places of birth, but generally they do not tell us anything about the recruitment and survival of offspring. Positioning data can document the long-distance dispersal of wolves (Ciucci et al. 2009), but they do not tell us anything about gene flow within the European metapopulation of wolves. GPS-positions can mark the end of life of an animal, but do not tell us anything about the cause of death. We need to combine GPS positioning data with field observations and surveys, population counts and genetics. Ultimately we need to correlate observed differences in movement behaviour and resource selection to fitness measures of the tagged individuals, e.g. body size, age, reproduction and longevity (Hebblewhite & Haydon 2010). AcknowledgementsOddgeir Andersen and an anonymous reviewer contributed strongly to an improved version of this manuscript. Many thanks to Jos Milner who improved the language, and to Harry Andreassen who encouraged me to write this review in the first place. ReferencesAmos, B. (2000) Lessons from the history of light microscopy. Nature Cell Biology, 2, E151. Anderson, C.R. & Lindzey, F.G. (2003) Estimating cougar predation rates from GPS location clusters. Journal of Wildlife Management, 67, 307-316. Avery, M.L., Humphrey, J.S., Daughtery, T.S., Fischer, J.W., Milleson, M.P., Tillman, E.A., Bruce, W.E. & Walter, W.D. (2011) Vulture flight behavior and implications for aircraft safety. Journal of Wildlife Management, 75, 1581-1587. Awkerman, J.A., Fukuda, A., Higuchi, H. & Anderson, D.J. (2005) Foraging activity and submesoscale habitat use of waved albatrosses Phoebastria irrorata during chick-brooding period. Marine Ecology-Progress Series, 291, 289-300. Bartlam-Brooks, H.L.A., Bonyongo, M.C. & Harris, S. (2011) Will reconnecting ecosystems allow long-distance mammal migrations to resume? A case study of a zebra Equus burchelli migration in Botswana. Oryx, 45, 210-216. Basille, M., Calenge, C., Marboutin, É., Andersen, R. & Gaillard, J.-M. (2008) Assessing habitat selection using multivariate statistics: Some refinements of the ecological-niche factor analysis. Ecological Modelling, 211, 233-240. Beckmann, J.P., Murray, K., Seidler, R.G. & Berger, J. (2012) Human-mediated shifts in animal habitat use: Sequential changes in pronghorn use of a natural gas field in Greater Yellowstone. Biological Conservation, 147, 222-233. Bengsen, A.J., Butler, J.A. & Masters, P. (2012) Applying home-range and landscape-use data to design effective feral-cat control programs. Wildlife Research, 39, 258-265. Bhatta, B. (2010) Global Navigation Satellite Systems : Insights into GPS, GLONASS, Galileo, Compass, and Others. BS Publications, Hyderabad, IND. Biggs, J.R., Bennett, K.D. & Fresquez, P.R. (2001) Relationship between home range characteristics and the probability of obtaining successful global positioning system (GPS) collar positions for elk in New Mexico. Western North American Naturalist, 61, 213-222. Bischof, R., Egil Loe, L., Meisingset, E.L., Zimmermann, B., Van Moorter, B. & Mysterud, A. (2012) A migratory northern ungulate in the pursuit of spring: jumping or surfing the green wave? American Naturalist, 180, 407. Bjørneraas, K., Van Moorter, B., Rolandsen, C.M. & Herfindal, I. (2010) Screening Global Positioning System location data for errors using animal movement characteristics. Journal of Wildlife Management, 74, 1361-1366. Blackie, H.M. (2010) Comparative performance of three brands of lightweight Global Positioning System collars. Journal of Wildlife Management, 74, 1911-1916. Blackie, H.M., Russell, J.C. & Clout, M.N. (2011) Maternal influence on philopatry and space use by juvenile brushtail possums (Trichosurus vulpecula). Journal of Animal Ecology, 80, 477-483. Blake, S., Deem, S.L., Mossimbo, E., Maisels, F. & Walsh, P. (2009) Forest elephants: Tree planters of the Congo. Biotropica, 41, 459-468. Blake, S., Douglas-Hamilton, I. & Karesh, W.B. (2001) GPS telemetry of forest elephants in Central Africa: results of a preliminary study. African Journal of Ecology, 39, 178-186. Blanc, F. & Brelurut, A. (1997) Short-term behavioral effects of equipping red deer hinds with a tracking collar. Zeitschrift Fur Saugetierkunde-International Journal of Mammalian Biology, 62, 18-26. Bohrer, G., Brandes, D., Mandel, J.T., Bildstein, K.L., Miller, T.A., Lanzone, M., Katzner, T., Maisonneuve, C. & Tremblay, J.A. (2012) Estimating updraft velocity components over large spatial scales: contrasting migration strategies of golden eagles and turkey vultures. Ecology Letters, 15, 96-103. Bowman, J.L., Kochanny, C.O., Demarais, S. & Leopold, B.D. (2000) Evaluation of a GPS collar for white-tailed deer. Wildlife Society Bulletin, 28, 141-145. Bradshaw, C.J.A., Sims, D.W. & Hays, G.C. (2007) Measurement error causes scale-dependent threshold erosion of biological signals in animal movement data. Ecological Applications, 17, 628-638. Brooks, C., Bonyongo, C. & Harris, S. (2008) Effects of global positioning system collar weight on zebra behavior and location error. Journal of Wildlife Management, 72, 527-534. Brooks, C.J. & Harris, S. (2008) Directed movement and orientation across a large natural landscape by zebras, Equus burchelli antiquorum. Animal Behaviour, 76, 277-285. Brooks, L.B., Harvey, J.T. & Nichols, W.J. (2009) Tidal movements of East Pacific green turtle Chelonia mydas at a foraging area in Baja California Sur, Mexico. Marine Ecology-Progress Series, 386, 263-274. Bunnefeld, N., Borger, L., van Moorter, B., Rolandsen, C.M., Dettki, H., Solberg, E.J. & Ericsson, G. (2011) A model-driven approach to quantify migration patterns: individual, regional and yearly differences. Journal of Animal Ecology, 80, 466-476. Cagnacci, F., Focardi, S., Heurich, M., Stache, A., Hewison, A.J.M., Morellet, N., Kjellander, P., Linnell, J.D.C., Mysterud, A., Neteler, M., Delucchi, L., Ossi, F. & Urbano, F. (2011) Partial migration in roe deer: migratory and resident tactics are end points of a behavioural gradient determined by ecological factors. Oikos, 120, 1790-1802. Cain, J.W., Krausman, P.R., Jansen, B.D. & Morgart, J.R. (2005) Influence of topography and GPS fix interval on GPS collar performance. Wildlife Society Bulletin, 33, 926-934. Calenge, C., Dray, S. & Royer-Carenzi, M. (2009) The concept of animals' trajectories from a data analysis perspective. Ecological Informatics, 4, 34-41. Campos-Arceiz, A., Larrinaga, A.R., Weerasinghe, U.R., Takatsuki, S., Pastorini, J., Leimgruber, P., Fernando, P. & Santamaria, L. (2008) Behavior rather than diet mediates seasonal differences in seed dispersal by asian elephants. Ecology, 89, 2684-2691. Cavalcanti, S.M.C. & Gese, E.M. (2009) Spatial ecology and social interactions of jaguars (Panthera onca) in the southern Pantanal, Brazil. Journal of Mammalogy, 90, 935-945. Cavalcanti, S.M.C. & Gese, E.M. (2010) Kill rates and predation patterns of jaguars (Panthera onca) in the southern Pantanal, Brazil. Journal of Mammalogy, 91, 722-736. Chetkiewicz, C.L.B. & Boyce, M.S. (2009) Use of resource selection functions to identify conservation corridors. Journal of Applied Ecology, 46, 1036-1047. Chivers, L.S., Lundy, M.G., Colhoun, K., Newton, S.F., Houghton, J.D.R. & Reid, N. (2012) Foraging trip time-activity budgets and reproductive success in the black-legged kittiwake. Marine Ecology-Progress Series, 456, 269-277. Ciucci, P., Reggioni, W., Maiorano, L. & Boitani, L. (2009) Long-distance dispersal of a rescued wolf from the northern Apennines to the western Alps. Journal of Wildlife Management, 73, 1300-1306. Colchero, F., Conde, D.A., Manterola, C., Chavez, C., Rivera, A. & Ceballos, G. (2011) Jaguars on the move: modeling movement to mitigate fragmentation from road expansion in the Mayan Forest. Animal Conservation, 14, 158-166. Conde, D.A., Colchero, F., Zarza, H., Christensen, N.L., Sexton, J.O., Manterola, C., Chavez, C., Rivera, A., Azuara, D. & Ceballos, G. (2010) Sex matters: Modeling male and female habitat differences for jaguar conservation. Biological Conservation, 143, 1980-1988. Cooper, R.M. & Millspaugh, J.J. (2001) Accounting for variation in resource availability and animal behavior in resource selection studies. Radio tracking and animal populations (eds J.J. Millspaugh & J.M. Marzluff), pp. 243-273. Academic Press, San Diego. Cooper, S.M., Perotto-Baldivieso, H.L., Owens, M.K., Meek, M.G. & Figueroa-Pagan, M. (2008) Distribution and interaction of white-tailed deer and cattle in a semi-arid grazing system. Agriculture Ecosystems & Environment, 127, 85-92. Cornelis, D., Benhamou, S., Janeau, G., Morellet, N., Ouedraogo, M. & de Visscher, M.N. (2011) Spatiotemporal dynamics of forage and water resources shape space use of West African savanna buffaloes. Journal of Mammalogy, 92, 1287-1297. Creel, S., Winnie, J., Maxwell, B., Hamlin, K. & Creel, M. (2005) Elk alter habitat selection as an antipredator response to wolves. Ecology, 86, 3387-3397. Cumming, G.S. & Ndlovu, M. (2011) Satellite telemetry of Afrotropical ducks: methodological details and assessment of success rates. African Zoology, 46, 425-434. D'Eon, R.G. & Delparte, D. (2005) Effects of radio-collar position and orientation on GPS radio-collar performance, and the implications of PDOP in data screening. Journal of Applied Ecology, 42, 383-388. D'Eon, R.G. & Serrouya, R. (2005) Mule deer seasonal movements and multiscale resource selection using global positioning system radiotelemetry. Journal of Mammalogy, 86, 736-744. D'Eon, R.G., Serrouya, R., Smith, G. & Kochanny, C.O. (2002) GPS radiotelemetry error and bias in mountainous terrain. Wildlife Society Bulletin, 30, 430-439. Dahl, F., Åhlén, P.A. & Granström, Å. (2010) The management of raccoon dogs (Nyctereutes procyonoides) in Scandinavia. Aliens, 30, 59-63. DeCesare, N.J., Squires, J.R. & Kolbe, J.A. (2005) Effect of forest canopy on GPS-based movement data. Wildlife Society Bulletin, 33, 935-941. Dell'Ariccia, G., Dell'Omo, G., Massa, B. & Bonadonna, F. (2010) First GPS-tracking of Cory's shearwater in the Mediterranean Sea. Italian Journal of Zoology, 77, 339-346. Dell'Ariccia, G., Dell'Omo, G., Wolfer, D.P. & Lipp, H.P. (2008) Flock flying improves pigeons' homing: GPS track analysis of individual flyers versus small groups. Animal Behaviour, 76, 1165-1172. Demma, D.J., Barber-Meyer, S.M. & Mech, L.D. (2007) Testing global positioning system telemetry to study wolf predation on deer fawns. Journal of Wildlife Management, 71, 2767-2775. Demma, D.J. & Mech, L.D. (2009) Wolf, Canis lupus, visits to white-tailed deer, Odocoileus virginianus, summer ranges: Optimal foraging? Canadian Field-Naturalist, 123, 299-303. Dennis, T.E., Rayner, M.J. & Walker, M.M. (2007) Evidence that pigeons orient to geomagnetic intensity during homing. Proceedings of the Royal Society B-Biological Sciences, 274, 1153-1158. Dettki, H. & Ericsson, G. (2008) Screening radiolocation datasets for movement strategies with time series segmentation. Journal of Wildlife Management, 72, 535-542. Di Orio, A.P., Callas, R. & Schaefer, R.J. (2003) Performance of two GPS telemetry collars under different habitat conditions. Wildlife Society Bulletin, 31, 372-379. Diggelen, F.v. (2009) A-GPS : Assisted GPS, GNSS, and SBAS. Artech House, Norwood, MA, USA. Dodd, N.L., Gagnon, J.W., Boe, S. & Schweinsburg, R.E. (2007) Assessment of elk highway permeability by using Global Positioning System telemetry. Journal of Wildlife Management, 71, 1107-1117. Dray, S., Royer-Carenzi, M. & Calenge, C. (2010) The exploratory analysis of autocorrelation in animal-movement studies. Ecological Research, 25, 673-681. Duchesne, T., Fortin, D. & Courbin, N. (2010) Mixed conditional logistic regression for habitat selection studies. Journal of Animal Ecology, 79, 548-555. Dussault, C., Courtois, R., Ouellet, J.P. & Huot, J. (1999) Evaluation of GPS telemetry collar performance for habitat studies in the boreal forest. Wildlife Society Bulletin, 27, 965-972. Dussault, C., Ouellet, J.P., Courtois, R., Huot, J., Breton, L. & Jolicoeur, H. (2005) Linking moose habitat selection to limiting factors. Ecography, 28, 619-628. Dussault, C., Ouellet, J.P., Courtois, R., Huot, J., Breton, L. & Larochelle, J. (2004) Behavioural responses of moose to thermal conditions in the boreal forest. Ecoscience, 11, 321-328. Dussault, C., Ouellet, J.P., Laurian, C., Courtois, R., Poulin, M. & Breton, L. (2007) Moose movement rates along highways and crossing probability models. Journal of Wildlife Management, 71, 2338-2345. Dyer, S.J., O'Neill, J.P., Wasel, S.M. & Boutin, S. (2001) Avoidance of industrial development by woodland caribou. Journal of Wildlife Management, 65, 531-542. Dyer, S.J., O'Neill, J.P., Wasel, S.M. & Boutin, S. (2002) Quantifying barrier effects of roads and seismic lines on movements of female woodland caribou in northeastern Alberta. Canadian Journal of Zoology-Revue Canadienne De Zoologie, 80, 839-845. Eriksen, A., Wabakken, P., Zimmermann, B., Andreassen, H.P., Arnemo, J.M., Gundersen, H., Liberg, O., Linnell, J., Milner, J.M., Pedersen, H.C., Sand, H., Solberg, E.J. & Storaas, T. (2011) Activity patterns of predator and prey: a simultaneous study of GPS-collared wolves and moose. Animal Behaviour, 81, 423-431. Eriksen, A., Wabakken, P., Zimmermann, B., Andreassen, H.P., Arnemo, J.M., Gundersen, H., Milner, J.M., Liberg, O., Linnell, J., Pedersen, H.C., Sand, H., Solberg, E.J. & Storaas, T. (2009) Encounter frequencies between GPS-collared wolves (Canis lupus) and moose (Alces alces) in a Scandinavian wolf territory. Ecological Research, 24, 547-557. Etherington, T.R., Pietravalle, S. & Cowan, D.P. (2010) Visualising uncertainty in radio-telemetry wildlife-tracking data to aid better study design. Wildlife Research, 37, 482-488. Evans, K., Baer, H., Bryant, E., Holland, M., Rupley, T. & Wilcox, C. (2011) Resolving estimation of movement in a vertically migrating pelagic fish: Does GPS provide a solution? Journal of Experimental Marine Biology and Ecology, 398, 9-17. Fieberg, J. & Kochanny, C.O. (2005) Quantifying home-range overlap: The importance of the utilization distribution. Journal of Wildlife Management, 69, 1346-1359. Fiedler, W. (2009) New technologies for monitoring bird migration and behaviour. Ringing & Migration, 24, 175-179. Fielitz, U., Renner, U., Schulte, R. & Wolfel, H. (1996) Satellite telemetry on red deer in the Harz - A pilot study. Zeitschrift Fur Jagdwissenschaft, 42, 1-11. Flesch, J.S., Duncan, M.G., Pascoe, J.H. & Mulley, R.C. (2009) A simple method of attaching GPS tracking devices to free-ranging lace monitors (Varanus varius). Herpetological Conservation and Biology, 4, 411-414. Fortin, D., Beyer, H.L., Boyce, M.S., Smith, D.W., Duchesne, T. & Mao, J.S. (2005) Wolves influence elk movements: Behavior shapes a trophic cascade in Yellowstone National Park. Ecology, 86, 1320-1330. Fortin, D., Fortin, M.E., Beyer, H.L., Duchesne, T., Courant, S. & Dancose, K. (2009) Group-size-mediated habitat selection and group fusion-fission dynamics of bison under predation risk. Ecology, 90, 2480-2490. Frair, J.L., Nielsen, S.E., Merrill, E.H., Lele, S.R., Boyce, M.S., Munro, R.H.M., Stenhouse, G.B. & Beyer, H.L. (2004) Removing GPS collar bias in habitat selection studies. Journal of Applied Ecology, 41, 201-212. Freitas, C., Kovacs, K.M., Andersen, M., Aars, J., Sandven, S., Skern-Mauritzen, M., Pavlova, O. & Lydersen, C. (2012) Importance of fast ice and glacier fronts for female polar bears and their cubs during spring in Svalbard, Norway. Marine Ecology-Progress Series, 447, 289-304. Fritz, H., Said, S. & Weimerskirch, H. (2003) Scale-dependent hierarchical adjustments of movement patterns in a long-range foraging seabird. Proceedings of the Royal Society B-Biological Sciences, 270, 1143-1148. Gagliardo, A., Savini, M., De Santis, A., Dell'Omo, G. & Ioale, P. (2009) Re-orientation in clock-shifted homing pigeons subjected to a magnetic disturbance: a study with GPS data loggers. Behavioral Ecology and Sociobiology, 64, 289-296. Galanti, V., Preatoni, D., Martinoti, A., Wauters, L.A. & Tosi, G. (2006) Space and habitat use of the African elephant in the Tarangire-Manyara ecosystem, Tanzania: Implications for conservation. Mammalian Biology, 71, 99-114. Garneau, D.E., Boudreau, T., Keech, M. & Post, E. (2008) Black bear movements and habitat use during a critical period for moose calves. Mammalian Biology, 73, 85-92. Gau, R.J., Mulders, R., Ciarniello, L.J., Heard, D.C., Chetkiewicz, C.L.B., Boyce, M., Munro, R., Stenhouse, G., Chruszcz, B., Gibeau, M.L., Milakovic, B. & Parker, K.L. (2004) Uncontrolled field performance of Televilt GPS-Simplex (TM) collars on grizzly bears in western and northern Canada. Wildlife Society Bulletin, 32, 693-701. Gervasi, V., Brunberg, S. & Swenson, J.E. (2006) An individual-based method to measure animal activity levels: A test on brown bears. Wildlife Society Bulletin, 34, 1314-1319. Girard, I., Dussault, C., Ouellet, J.P., Courtois, R. & Caron, A. (2006) Balancing number of locations with number of individuals in telemetry studies. Journal of Wildlife Management, 70, 1249-1256. Gradev, G.G., Matarranz, V.G., Dobreva, E.H., Popov, D.V., Ivanov, II, Klisurov, II, Kmetova, E.V. & Vasilakis, D.P. (2011) First results of the tracking of an eastern imperial eagle (Aquila heliaca) tagged with a GPS/GSM transmitter in Bulgaria. Acta Zoologica Bulgarica, 15-20. Graham, M.D., Douglas-Hamilton, I., Adams, W.M. & Lee, P.C. (2009) The movement of African elephants in a human-dominated land-use mosaic. Animal Conservation, 12, 445-455. Gremillet, D., Dell'Omo, G., Ryan, P.G., Peters, G., Ropert-Coudert, Y. & Weeks, S.J. (2004) Offshore diplomacy, or how seabirds mitigate intra-specific competition: a case study based on GPS tracking of Cape gannets from neighbouring colonies. Marine Ecology-Progress Series, 268, 265-279. Grosman, P.D., Jaeger, J.A.G., Biron, P.M., Dussault, C. & Ouellet, J.P. (2009) Reducing moose-vehicle collisions through salt pool removal and displacement: an agent-based modeling approach. Ecology and Society, 14. Grosman, P.D., Jaeger, J.A.G., Biron, P.M., Dussault, C. & Ouellet, J.P. (2011) Trade-off between road avoidance and attraction by roadside salt pools in moose: An agent-based model to assess measures for reducing moose-vehicle collisions. Ecological Modelling, 222, 1423-1435. Gustine, D.D. & Parker, K.L. (2008) Variation in the seasonal selection of resources by woodland caribou in northern British Columbia. Canadian Journal of Zoology-Revue Canadienne De Zoologie, 86, 812-825. Guthrie, J.D., Byrne, M.E., Hardin, J.B., Kochanny, C.O., Skow, K.L., Snelgrove, R.T., Butler, M.J., Peterson, M.J., Chamberlain, M.J. & Collier, B.A. (2011) Evaluation of a global positioning system backpack transmitter for wild turkey research. Journal of Wildlife Management, 75, 539-547. Habib, T.J., Merrill, E.H., Pybus, M.J. & Coltman, D.W. (2011) Modelling landscape effects on density-contact rate relationships of deer in eastern Alberta: Implications for chronic wasting disease. Ecological Modelling, 222, 2722-2732. Haddadi, H., King, A.J., Wills, A.P., Fay, D., Lowe, J., Morton, A.J., Hailes, S. & Wilson, A.M. (2011) Determining association networks in social animals: choosing spatial-temporal criteria and sampling rates. Behavioral Ecology and Sociobiology, 65, 1659-1668. Haines, A.M., Grassman, L.I., Tewes, M.E. & Janecka, J.E. (2006) First ocelot (Leopardus pardalis) monitored with GPS telemetry. European Journal of Wildlife Research, 52, 216-218. Hamer, K.C., Humphreys, E.M., Garthe, S., Hennicke, J., Peters, G., Gremillet, D., Phillips, R.A., Harris, M.P. & Wanless, S. (2007) Annual variation in diets, feeding locations and foraging behaviour of gannets in the North Sea: flexibility, consistency and constraint. Marine Ecology-Progress Series, 338, 295-305. Haramis, G.M. & White, T.S. (2011) A beaded collar for dual micro GPS/VHF transmitter attachment to nutria. Mammalia, 75, 79-82. Hayward, M.W., Hayward, G.J., Druce, D.J. & Kerley, G.I.H. (2009) Do fences constrain predator movements on an evolutionary scale? Home range, food intake and movement patterns of large predators reintroduced to Addo Elephant National Park, South Africa. Biodiversity and Conservation, 18, 887-904. Hazel, J. (2009) Evaluation of fast-acquisition GPS in stationary tests and fine-scale tracking of green turtles. Journal of Experimental Marine Biology and Ecology, 374, 58-68. Hebblewhite, M. & Haydon, D.T. (2010) Distinguishing technology from biology: a critical review of the use of GPS telemetry data in ecology. Philosophical Transactions of the Royal Society B-Biological Sciences, 365, 2303-2312. Hebblewhite, M. & Merrill, E.H. (2007) Multiscale wolf predation risk for elk: does migration reduce risk? Oecologia, 152, 377-387. Hemson, G., Johnson, P., South, A., Kenward, R., Ripley, R. & Macdonald, D. (2005) Are kernels the mustard? Data from global positioning system (GPS) collars suggests problems for kernel home-range analyses with least-squares cross-validation. Journal of Animal Ecology, 74, 455-463. Heurich, M., Traube, M., Stache, A. & Lottker, P. (2012) Calibration of remotely collected acceleration data with behavioral observations of roe deer (Capreolus capreolus L.). Acta Theriologica, 57, 251-255. Horne, J.S., Garton, E.O. & Sager-Fradkin, K.A. (2007) Correcting home-range models for observation bias. Journal of Wildlife Management, 71, 996-1001. Humle, T., Colin, C., Laurans, M. & Raballand, E. (2011) Group release of sanctuary chimpanzees (Pan troglodytes) in the Haut Niger National Park, Guinea, West Africa: Ranging patterns and lessons so far. International Journal of Primatology, 32, 456-473. Inman, R.M., Packila, M.L., Inman, K.H., McCue, A.J., White, G.C., Persson, J., Aber, B.C., Orme, M.L., Alt, K.L., Cain, S.L., Fredrick, J.A., Oakleaf, B.J. & Sartorius, S.S. (2012) Spatial ecology of wolverines at the southern periphery of distribution. Journal of Wildlife Management, 76, 778-792. Johnson, D.H. (1980) The comparison of usage and availability measurements for evaluating resource preference. Ecology, 61, 65-71. Kaplan, E.D. & Hegarty, C.J. (2006) Understanding GPS: principles and applications, 2 edn. Artech House, Boston, London. Kays, R., Jansen, P.A., Knecht, E.M.H., Vohwinkel, R. & Wikelski, M. (2011) The effect of feeding time on dispersal of Virola seeds by toucans determined from GPS tracking and accelerometers. Acta Oecologica-International Journal of Ecology, 37, 625-631. Kenward, R. (2001) Historical and practical perspectives. Radio tracking and animal populations (eds J.J. Millspaugh & J.M. Marzluff), pp. 3-12. Academic Press, San Diego. Kenward, R.E. (2000) A manual for wildlife radio tagging. Academic Press. Kranstauber, B., Kays, R., LaPoint, S.D., Wikelski, M. & Safi, K. (2012) A dynamic Brownian bridge movement model to estimate utilization distributions for heterogeneous animal movement. Journal of Animal Ecology, 81, 738-746. Lele, S.R. & Keim, J.L. (2006) Weighted distributions and estimation of resource selection probability functions. Ecology, 87, 3021-3028. Macdonald, D.W. (1978) Radio-tracking: some applications and limitations. Animal marking: Recognition marking of animals in research. (ed. G.B. Stenhouse), pp. 192-204. Macmillan, London. Meade, J., Biro, D. & Guilford, T. (2005) Homing pigeons develop local route stereotypy. Proceedings of the Royal Society B-Biological Sciences, 272, 17-23. Millspaugh, J.J. & Marzluff, J.M. (2001) Radio tracking and animal populations. Academic Press, San Diego. Moen, R., Pastor, J., Cohen, Y. & Schwartz, C.C. (1996) Effects of moose movement and habitat use on GPS collar performance. Journal of Wildlife Management, 60, 659-668. Morales, J.M., Moorcroft, P.R., Matthiopoulos, J., Frair, J.L., Kie, J.G., Powell, R.A., Merrill, E.H. & Haydon, D.T. (2010) Building the bridge between animal movement and population dynamics. Philosophical Transactions of the Royal Society B: Biological Sciences, 365, 2289-2301. Morehouse, A.T. & Boyce, M.S. (2011) From venison to beef: seasonal changes in wolf diet composition in a livestock grazing landscape. Frontiers in Ecology and the Environment, 9, 440-445. Neumann, W., Ericsson, G. & Dettki, H. (2010) Does off-trail backcountry skiing disturb moose? European Journal of Wildlife Research, 56, 513-518. Rempel, R.S., Rodgers, A.R. & Abraham, K.F. (1995) Performance of a GPS animal location system under boreal forest canopy. Journal of Wildlife Management, 59, 543-551. Rodgers, A.R. (2001) Recent telemetry technology. Radio tracking and animal populations (eds J.J. Millspaugh & J.M. Marzluff), pp. 79-121. Academic Press, San Diego. Rodgers, A.R., Rempel, R.S. & Abraham, K.F. (1996) A GPS-based telemetry system. Wildlife Society Bulletin, 24, 559-566. Sundell, J., Kojola, I. & Hanski, I. (2006) A new GPS-GSM-based method to study behavior of brown bears. Wildlife Society Bulletin, 34, 446-450. Theiss, A., Yen, D.C. & Ku, C.-Y. (2005) Global Positioning Systems: an analysis of applications, current development and future implementations. Computer Standards & Interfaces, 27, 89-100. Tomkiewicz, S.M., Fuller, M.R., Kie, J.G. & Bates, K.K. (2010) Global positioning system and associated technologies in animal behaviour and ecological research. Philosophical Transactions of the Royal Society B: Biological Sciences, 365, 2163-2176. Withey, J.C., Bloxton, T.D. & Marzluff, J.M. (2001) Effects of tagging and location error in wildlife radiotelemetry studies. Radio tracking and animal populations (eds J.J. Millspaugh & J.M. Marzluff), pp. 45-69. Academic Press, San Diego. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||